

中國科學(xué)院南海海洋研究所熱帶海洋生物資源與生態(tài)重點(diǎn)實(shí)驗(yàn)室研究員高貝樂團(tuán)隊(duì)以具有足夠進(jìn)化深度和生態(tài)多樣性的彎曲菌門為研究對象,分析了六大信號轉(zhuǎn)導(dǎo)系統(tǒng)在該門不同分支的進(jìn)化過程及復(fù)雜度構(gòu)建的方式,解析了細(xì)菌的信號網(wǎng)絡(luò)從簡單演變?yōu)閺?fù)雜,或從復(fù)雜至簡單的過程。相關(guān)研究成果近日以“Evolutionary Principles of Bacterial Signaling Capacity and Complexity”為題發(fā)表于mBio。
細(xì)菌依賴信號轉(zhuǎn)導(dǎo)系統(tǒng)來感知和響應(yīng)環(huán)境變化以維持生存和繁殖,目前已知的細(xì)菌信號轉(zhuǎn)導(dǎo)系統(tǒng)主要有六類:1.趨化系統(tǒng),主要指揮細(xì)菌的運(yùn)動方向; 2.雙組份信號轉(zhuǎn)導(dǎo)系統(tǒng),調(diào)節(jié)基因的轉(zhuǎn)錄與表達(dá);3.第二信使介導(dǎo)的信號轉(zhuǎn)導(dǎo)系統(tǒng),例如環(huán)化腺苷酸、環(huán)二鳥苷酸;4.STYK磷酸激酶和磷酸酶介導(dǎo)的信號通路;5.ECF介導(dǎo)的信號通路,調(diào)節(jié)基因的轉(zhuǎn)錄和表達(dá);6.用于細(xì)菌間交流的群體感應(yīng)。這些信號轉(zhuǎn)導(dǎo)系統(tǒng)可以獨(dú)立存在,也可以交互在一起,形成一個(gè)網(wǎng)絡(luò)。以往的調(diào)查研究對不同細(xì)菌物種中的幾個(gè)信號轉(zhuǎn)導(dǎo)系統(tǒng)的組成蛋白進(jìn)行了統(tǒng)計(jì)和分類,以了解它們的系統(tǒng)組成和多樣性、模塊化和可塑性、系統(tǒng)發(fā)育分布、與基因組大小和生態(tài)分布的相關(guān)性等等。然而,目前尚未完全清楚細(xì)菌的信號轉(zhuǎn)導(dǎo)網(wǎng)絡(luò)在整體上是如何演變的。
回答該問題,需要一個(gè)具有足夠進(jìn)化深度和生態(tài)多樣性的細(xì)菌門作為分析框架。系統(tǒng)發(fā)育基因組學(xué)分析表明,彎曲菌門最早期的進(jìn)化分支由棲息于深海熱液口的嚴(yán)格厭氧和嗜熱的化能自養(yǎng)菌組成,該環(huán)境被認(rèn)為是太古宙地球環(huán)境的模式系統(tǒng),表明彎曲菌門的早期進(jìn)化分支在早期地球歷史中發(fā)揮了重要作用。隨后的進(jìn)化分支向海洋和陸地淡水系統(tǒng)進(jìn)行生態(tài)位擴(kuò)張。隨著生態(tài)環(huán)境的多樣化,這些譜系表現(xiàn)出從厭氧到微氧/好氧,從嗜熱到嗜中溫,以及從自養(yǎng)到異養(yǎng)的轉(zhuǎn)變。其中一些譜系成為與宿主相關(guān)的共生菌或病原菌,以嚴(yán)格嗜中溫和異養(yǎng)型的Campylobacter屬和Helicobacter屬為代表。此外,該門物種的基因組大小變化范圍較窄(1.4-3.5Mb),沒有發(fā)生劇烈的基因組縮減或擴(kuò)張。顯然,彎曲菌門是一個(gè)具有足夠進(jìn)化深度、分支次序有據(jù)、生態(tài)分布廣泛的單系群,為研究細(xì)菌信號轉(zhuǎn)導(dǎo)系統(tǒng)或信號網(wǎng)絡(luò)的進(jìn)化提供了一個(gè)很好的“生態(tài)-進(jìn)化”框架(圖1)。
研究人員利用彎曲菌門的“生態(tài)-進(jìn)化”框架,對門下物種的82和基因組進(jìn)行了詳細(xì)的比較基因組學(xué)分析。研究發(fā)現(xiàn),整體上彎曲菌門物種的信號轉(zhuǎn)導(dǎo)網(wǎng)絡(luò)的進(jìn)化模式表現(xiàn)出兩種相反的趨勢。在生態(tài)位擴(kuò)張的物種中,信號網(wǎng)絡(luò)的復(fù)雜性隨著基因家族的擴(kuò)張而增加,水平基因轉(zhuǎn)移、基因復(fù)制、融合和裂變等新基因獲得方式為基因創(chuàng)新和不同轉(zhuǎn)導(dǎo)機(jī)制的整合創(chuàng)造了機(jī)會。相反,當(dāng)細(xì)菌分支專一性適應(yīng)特殊生態(tài)位時(shí),其信號網(wǎng)絡(luò)的復(fù)雜性隨著大量基因的丟失而降低。具體到每個(gè)信號轉(zhuǎn)導(dǎo)系統(tǒng),我們發(fā)現(xiàn)轉(zhuǎn)導(dǎo)機(jī)制的不同導(dǎo)致進(jìn)化方式也有差別。主要結(jié)論包括:1.趨化系統(tǒng)主要通過水平基因轉(zhuǎn)移獲得新的整套趨化基因簇來增加復(fù)雜度,而不同趨化類別之間極少發(fā)生融合;2.雙組份系統(tǒng)的復(fù)雜度主要取決于非經(jīng)典組氨酸激酶的數(shù)量,在經(jīng)典組氨酸激酶的結(jié)構(gòu)域組成的基礎(chǔ)上通過在激酶的N-端或者C-端融合REC結(jié)構(gòu)域,來實(shí)現(xiàn)磷酸信號的多步傳遞、輸入和輸出信號的整合或分流;3.環(huán)二鳥苷酸介導(dǎo)的通路進(jìn)化方式最多樣,其合成酶和分解酶容易獲得也容易丟失,因?yàn)榈诙攀剐》肿拥暮铣珊徒到獬杀镜停以撔》肿硬皇艿鞍谆プ鞯倪M(jìn)化約束。以上三種系統(tǒng)在彎曲菌門的祖先細(xì)胞中就已經(jīng)進(jìn)化了,后來的分支隨著生態(tài)位的拓張不斷增加網(wǎng)絡(luò)復(fù)雜度;而宿主體內(nèi)的共生菌和病原菌則大量丟失信號轉(zhuǎn)導(dǎo)蛋白,只保留簡單的單組份、雙組份系統(tǒng)以及與鞭毛共進(jìn)化的趨化系統(tǒng),一些分支徹底丟了環(huán)二鳥苷酸介導(dǎo)的通路。總而言之,在彎曲菌門中,信號轉(zhuǎn)導(dǎo)網(wǎng)絡(luò)的大小和復(fù)雜度呈現(xiàn)共同增強(qiáng)與共同減弱的趨勢,由細(xì)菌感受環(huán)境變化的需求、能量成本、遺傳資源和基因組內(nèi)的協(xié)同進(jìn)化共同決定的。
中國科學(xué)院南海海洋研究所2022屆博士莫然、2020級博士生陳園園、2020屆碩士劉煜耿為該論文的共同第一作者,高貝樂研究員為通訊作者。本研究得到了國家自然科學(xué)基金、南方海洋科學(xué)與工程引進(jìn)人才團(tuán)隊(duì)重點(diǎn)專項(xiàng)廣東實(shí)驗(yàn)室(廣州)、中國科學(xué)院戰(zhàn)略性先導(dǎo)專項(xiàng)、中國科學(xué)院南海生態(tài)與環(huán)境工程創(chuàng)新研究院的資助。
相關(guān)論文信息:https://journals.asm.org/doi/10.1128/mbio.00764-22
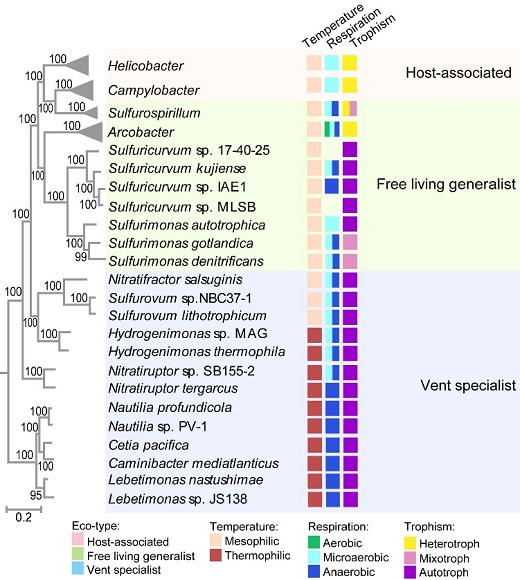
圖1 彎曲菌門的物種進(jìn)化樹與生態(tài)生理特點(diǎn)
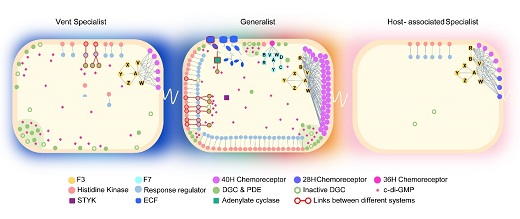
圖2 彎曲菌門代表物種的信號轉(zhuǎn)導(dǎo)網(wǎng)絡(luò)的構(gòu)成
附件下載: